| Gli
oftalmofrattali Alfredo Parrozzani |
www.oculistanet.it |
PARTE SECONDA
(continuazione)
9 - IL CORPO CILIARE
Il corpo ciliare è diviso in due parti: la Parte Plicata
e la Parte Piana. La prima è caratterizzata da 70-80
pliche o processi radiali, che a loro volta sono formati
da numerose altre pieghe più piccole con direzione
diversa. La parte piana è una regione appiattita che si
estende posteriormente, dalla parte plicata all'ora
serrata della retina.
La presenza di pieghe nella parte plicata , simili a
scale diverse, fa pensare ad una autosomiglianza. Pieghe
radiali composte da altre pieghe disposte in direzioni
diverse vengono a formare una struttura frattale.
La superficie risultante del corpo ciliare, formata da
pieghe su pieghe, è molto estesa. Questa struttura molto
irregolare consente di mantenere elevato il rapporto
superficie/volume, necessario per gli scambi di gas, di
liquidi e di elettroliti.
Il profilo della parte plicata del corpo ciliare è molto
simile ad una di quelle "curve mostro" frattali
originate da modificazioni iterative su una figura
geometrica di partenza, come la curva di Koch (realizzata
partendo da un triangolo equilatero, modificando i lati
con un altro triangolo fino ad ottenere una forma
stellata o a fiocco di neve).
Partendo da un esagono o da un dodecagono, eseguendo le
stesse modificazioni iterative dei lati come nella curva
di Koch (si modificano i lati con una forma a triangolo
il cui lato è 1/3 del lato di partenza, e si ripete
l'azione nei lati delle figure risultanti), si ottiene un
profilo irregolare con perimetro simile a quello della
parte plicata.
Il frattale che incontriamo in natura nel corpo ciliare
non possiede la regolarità di una curva di Koché ma è
autosimile in modo statistico. Le "penisole" e
le "baie" di una curva di Koch assomigliano
straordinariamente al profilo della parte plicata, e
anche se le protuberanze e le anse di questa non
ricorrono con la stessa successione a tutte le scale,
l'aspetto generale finisce per avere lo stesso aspetto
plicato a vari ingrandimenti.
Un'altra significativa differenza tra la curva di Koch e
un profilo del corpo ciliare sta nel fatto che la curva
è una forma matematica idealizzata che possiede una
struttura a infiniti livelli, mentre il secondo non è
una linea infinitamente complessa. Tuttavia con un
frattale si ha un modello migliore, anche se non
necessariamente perfetto, di una forma classica più
regolare e di complessità finita.
Con un frattale, oltre all'utilizzazione del modello
anatomico a più livelli, è possibile dare delle
interpretazioni fisiologiche e logiche al rapporto
forma-funzione, senza accettare passivamente i rilievi
morfologici.
Perché questa complessità frattale del corpo ciliare?
Probabilmente è necessaria una struttura così complessa
con elevata superficie di contatto con i liquidi
intraoculari per permettere una sufficiente e continua
produzione di acqueo, con scambio chimico-fisico di acqua
ed elettroliti.
Inoltre la stessa struttura poliplicata è necessaria per
una distribuzione delle forze di trazione continua delle
fibre zonulari nei movimenti accomodativi e di
sospensione meccanica del cristallino.
10 - L'IDRODINAMICA OCULARE
La pressione oculare normalmente si aggira sui 12-18
mmHg, tale pressione può essere considerata la
risultante di due meccanismi principali: la produzione e
il deflusso dell'umor acqueo. Un costante equilibrio tra
queste due azioni consente alla pressione oculare di
conservare un suo valore caratteristico, considerato
normale, nelle 24 ore. Questo valore è individuale,
tuttavia non deve superare una certa soglia, oltre la
quale vengono a determinarsi danni anatomo-funzionali
irreversibili.
Molte condizioni oculari possono alterare il suddetto
equilibrio, modificando i valori pressori. In questa sede
non è opportuno prolungarsi su quali sono le situazioni
patologiche e quali i danni risultanti. Le variazioni
della pressione oculare sono legate anche a fattori di
natura sistemica, che possono provocare fluttuazioni dei
valori pressori. Le oscillazioni normali non superano i
2-3 mmHg, espressione di un controllo omeostatico da
parte dei parametri che regolano l'idrodinamica oculare.
Le fluttuazioni del tono oculare possono essere
interpretate in termini di omeostasi: i sistemi biologici
normalmente si comportano in modo da ridurre la
variabilità e mantenere costanti le funzioni interne.
Qualsiasi variabile fisiologica, incluso il tono oculare,
dovrebbe ritornare al suo stato stazionario dopo essere
stata perturbata. Il principio dell'omeostasi suggerisce
che le variazioni pressorie oculari sono semplicemente
risposte transitorie a un ambiente oscillante.
Si potrebbe immaginare che, in condizioni patologiche,
l'organismo perda la capacità di mantenere un valore
pressorio abbastanza costante e che quindi aumentino la
variabilità e le oscillazioni. Ma le cose si presentano
in modo diverso a chi misuri scrupolosamente i valori
pressori in tutto l'arco della giornata, e a varie scale
temporali.
La successione temporale ottenuta sembra irregolare e
completamente casuale, a prima vista; i valori pressori
sembrano non avere alcuna logica di distribuzione, a
parte un lieve ritmo giornaliero.
Esaminando però il grafico a differenti scale temporali,
è possibile rilevare alcune caratteristiche. Se si
osserva la serie temporale relativa all'arco di poche
ore, si riscontrano fluttuazioni , le cui caratteristiche
e la cui sequenza sembrano simili a quelle trovate in
serie temporali più lunghe. A scale temporali minori
(minuti), è possibile osservare fluttuazioni minori che
sembrano sempre molto simili all'andamento iniziale.
Le fluttuazioni tonometriche in scale temporali
differenti appaiono autosimili, come le ramificazioni di
un frattale geometrico. Questo risultato ci permette di
pensare che i meccanismi di controllo del tono oculare
possano essere di tipo caotico.
In altri termini i meccanismi che influiscono
sull'idrodinamica oculare sono tanti, molteplici ed
imprevedibili. Poiché in questi fenomeni non esiste una
chiara relazione tra causa ed effetto, si dice che
contengono elementi aleatori.
Una volta si riteneva che bastasse raccogliere ed
elaborare una quantità sufficiente di informazioni per
poter studiare e controllare completamente l'idrodinamica
oculare. In sistemi come questo l'aleatorietà non
scompare, anche se si raccolgono ulteriori informazioni.
A questo genere di aleatorietà si è dato il nome di
caos. Un paradosso apparente è che il caos è
deterministico, cioè è generato da regole fisse che, di
per s‚ non contengono alcun elemento casuale.
Il futuro immediato della regolazione dell'idrodinamica
oculare è determinato completamente dalle azioni
eseguite nel presente, tuttavia piccole indeterminazioni
vengono amplificate nel futuro lontano; quindi il
comportamento, benché prevedibile a breve scadenza, a
lungo termine risulta totalmente imprevedibile.
Sappiamo che un farmaco ipotonizzante è efficace perché
otteniamo un immediato controllo tonometrico, tuttavia a
distanza tale risultato può non essere sufficiente
perché intervengono altri fattori, seppur minimi, a
modificare decisamente l'equilibrio.
Perché un individuo con tono superiore alla media
considerata normale non sviluppa danni campimetrici,
mentre altri con uguale tono in poco tempo mostrano una
sofferenza glaucomatosa ed alterazioni campimetriche ?
Perché altri con tono considerato normale hanno un
deficit campimetrico glaucomatoso, che noi abbiamo
etichettato arbitrariamente "glaucoma a bassa
tensione", e rispondono poco alla terapia
ipotonizzante? perché spesso il danno obiettivo del
nervo ottico non evolve parallelamente al deficit
glaucomatoso del campo visivo? Tutte queste domande, e
tantissime altre, rischiano di restare senza risposta se
non consideriamo la natura intrinsecamente caotica
dell'idrodinamica oculare. In essa intervengono numerosi
fattori locali, sistemici ed esterni, che seppure
riuscissimo a conoscerli tutti, misurarli ed elaborarli,
non riusciremo mai a giungere ad una prevedibilità
rigorosa, perché persisterà sempre una certa
aleatorietà essenziale per la natura caotica del
sistema.
Per analizzare questo tipo di sistema dinamico complesso
non lineare si può ricorrere alla rappresentazione nello
spazio delle fasi o spazio degli stati. Questa tecnica
segue i valori delle variabili indipendenti che cambiano
nel tempo.
Le numerose variabili indipendenti presenti in molti
sistemi complessi sono spesso non identificabili e
misurabili. Tali sistemi possono essere rappresentati
nello spazio delle fasi con il metodo delle "delay
maps". L'ascissa di ogni punto corrisponde al valore
di una variabile in un certo istante, mentre l'ordinata
rappresenta il valore della stessa variabile dopo un
ritardo costante. L'insieme di questi punti in tempi
successivi forma una curva o traiettoria che descrive
l'evoluzione del sistema.
Per identificare il tipo di sistema dinamico si cerca un
attrattore della curva, cioè la regione di spazio delle
fasi che attrae le traiettorie.
L'attrattore può essere di tre tipi:
1) Un punto fisso, o un'area
molto limitata da essere sovrapponibile a un punto. Esso
descrive un sistema che evolve verso un singolo stato,
per esempio un pendolo smorzato. Nello spazio delle fasi
tutte le traiettorie convergono verso quel punto
attrattore.
2) Il ciclo limite, è un sistema che evolve verso uno
stato periodico, come un pendolo ideale senza attriti.
3) Attrattori strani, sono altri attrattori che
descrivono sistemi che non presentano caratteristiche
periodiche e non evolvono verso un punto fisso. Due
traiettorie che presentano condizioni iniziali
praticamente identiche si allontanano rapidamente e
diventano molto differenti, su distanze temporali
maggiori. Un sistema di questo tipo con attrattori strani
è caotico.
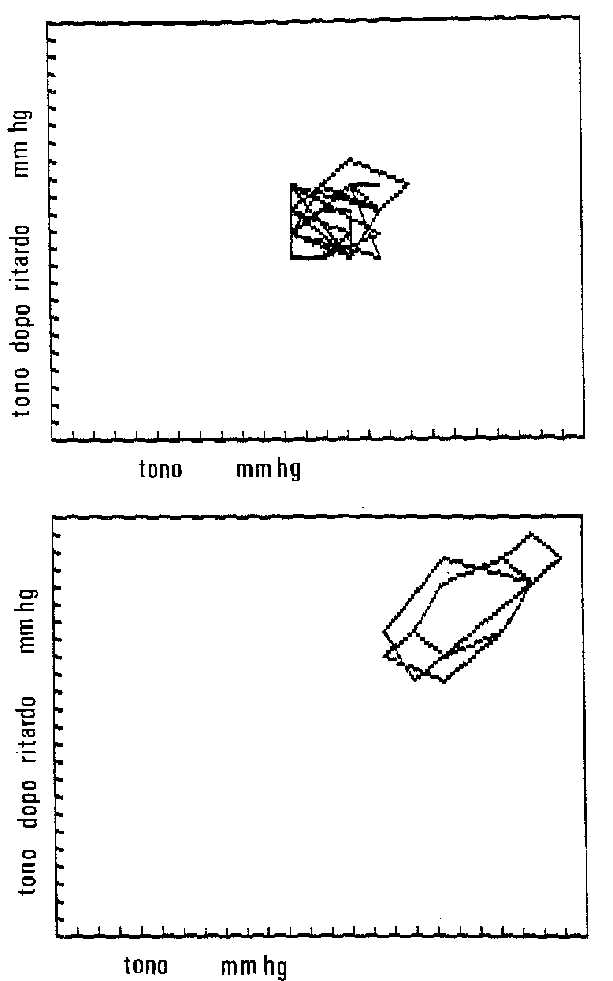
Ho analizzato la rappresentazione dello spazio delle fasi
per il tono oculare in rapporto al tempo, nell'occhio
normale e in situazioni di patologia dell'idrodinamica
oculare.
I risultati mostrano un comportamento simile ad un
attrattore strano nell'occhio normale, espressione di un
comportamento caotico del tono, seppur entro certi limiti
di valore tonometrico.
Perché questo comportamento? La idrodinamica caotica
consente di avere una risposta idonea alle influenze
dall'ambiente e ai fattori endogeni.
In situazioni di patologia dell'idrodinamica l'attrattore
presenta un aspetto talvolta più periodico, tipo ciclo
limite, seppur talvolta con note di comportamento
caotico,come nel glaucoma cronico.
Altre volte l'attrattore può essere di tipo a punto
fisso, come nel caso di glaucoma a chiusura d'angolo .
In entrambi i casi viene a mancare la caratteristica
caotica "fisiologica" di un sistema adattabile
e flessibile, compare l'aspetto periodico e rigido di un
sistema "patologico".
Questa caratteristica caotica di un sistema fisiologico
dipende dalle numerose variabili e fattori interni ed
esterni che entrano in gioco. In condizioni patologiche
il sistema può risentire di pochi fattori, cioè quelli
che prendono il sopravvento nell'equilibrio caotico
determinando la situazione patologica, per cui
l'attrattore prende l'aspetto periodico.
In situazioni drammatiche in cui l'equilibrio
dell'idrodinamica è notevolmente compromesso, come in
caso di chiusura d'angolo, l'attrattore assume un aspetto
quasi di punto fisso; questo dipende dal prevalere di un
solo fattore che condiziona totalmente tutto il sistema,
allontanandolo dalla dinamica caotica.
Questo nuovo modello dell'idrodinamica oculare da una
parte comporta l'esistenza di nuove limitazioni alla
capacità di compiere previsioni sul decorso e sulla
prognosi del sistema idrodinamico oculare fisiologico o
patologico; dall'altra , il determinismo inerente al caos
implica che molti fenomeni aleatori sono più prevedibili
di quanto si pensasse. Informazioni apparentemente
aleatorie possono essere spiegate in termini di leggi
semplici.
Con una interpretazione frattale dell'idrodinamica
oculare, soggetta alla legge del caos, l'impostazione
terapeutica di un'alterazione della stessa dovrebbe
essere modificata.
L'uso dei farmaci nel controllo dell'idrodinamica oculare
spesso può essere considerato un fallimento. Se non si
raggiunge un compenso sufficiente le conseguenze più
drammatiche non possono essere evitate a lungo termine.
A mio giudizio i metodi tradizionali per trattare questo
sistema idrodinamico estremamente instabile e complesso
sono troppo lineari e riduzionistici.
Il paradigma "ipertono --> patologia glaucomatosa
--> danno funzionale" è praticamente
intoccabile, anche se spesso avviene che il danno
funzionale progredisce anche ad avvenuto compenso
quantitativo del tono; allora si "inventa" il
glaucoma a bassa tensione o altre sindromi
"anomale", non essendo più in grado di
spiegare il fallimento di una terapia così lineare.
Non dobbiamo più raggiungere un semplice compenso
tonometrico, rapportato e collegato all'evoluzione del
campo visivo e dell'escavazione del disco ottico, ma
sarà necessario riportare il sistema idrodinamico
alterato, rigido, ad una dinamica caotica più adattabile
e flessibile, che consente all'occhio di rispondere alle
variazioni interne ed esterne, in tal modo le strutture
più sensibili saranno al riparo dai danni.
Il farmaco, o il trattamento chirurgico o parachirugico,
ideale non deve ridurre il tono oculare linearmente,
anche se drasticamente, ma deve ripristinare e rispettare
l'andamento caotico del sistema , anche se i valori
tonometrici risultanti non sono sempre inferiori al
valore soglia considerato normale.
11 - IL CRISTALLINO
La sostanza propria del cristallino è formata da fibre
notevolmente allungate, dirette in senso meridiano. I
margini di queste fibre si presentano sempre più
irregolari man mano che ci si avvicina al centro del
cristallino. Si possono notare, al microscopio
elettronico a scansione, creste, nicchie e dentellature.
Queste irregolarità sono di forma varia, con ulteriori
irregolarità a scala minore. La disposizione di queste
non presenta una regolarità, ma una disposizione
casuale. In tal modo si viene a creare una struttura
frattale con autosomiglianza statistica a più scale.
Questa irregolarità frattale delle fibre è necessaria
per creare una intima e solida unione tra le stesse.
Le proteine contenute nel cristallino normale si
presentano come molecole sferiche isolate. In condizioni
patologiche, determinate da vari fattori, queste molecole
sferiche mostrano fenomeni di aggregazione con comparsa
di aggregati giganteschi, la cui configurazione ha un
aspetto di aggregazione per diffusione.
Immaginiamo di far crescere un aggregato di particelle
aggiungendone una alla volta, in modo che appena viene a
contatto con l'oggetto in accrescimento vi aderisca.
Questo processo viene detto aggregazione. Quando
l'aggregato è piccolo più particelle aderiscono per
puro caso in una certa regione, formando piccole
protuberanze.
Le nuove particelle aderiranno più facilmente sulle
protuberanze, determinando un accrescimento delle stesse.
Alla fine della crescita la suddivisione delle
protuberanze dà origine a un frattale.
E' possibile far crescere aggregati simili anche su un
personal computer, creando modelli frattali realizzati
per aggregazione per diffusione simulata.
L'aggregazione per diffusione è una buona idealizzazione
di ciò che accade alle proteine sferiche del
cristallino. Queste proteine denaturate, da agenti
endogeni o esogeni, aggregandosi realizzano grosse
formazioni di aspetto frattale, che possono a loro volta
riunirsi per formare ammassi più grossolani di varia
dimensione.
Questi aggregati determinano una opacizzazione del
cristallino per perdita delle sue proprietà di
trasparenza.
Le opacità risultanti, anche se con forma e topografia
variabile in base agli agenti eziopatogenetici,
presentano una distribuzione irregolare legata al caso
con una omotetia interna. Questa autosomiglianza fa sì
che il caso abbia la stessa importanza a più scale nella
distribuzione delle opacità.
12 - LA CIRCOLAZIONE RETINICA
L'arteria centrale della retina, vera arteria e non
arteriola, entra nel bulbo penetrando nel nervo ottico e
fuoriuscendo da questo si suddivide nei suoi rami
secondari e terziari dell'arteria centrale della retina,
irrorando i quattro quadranti della retina. Le successive
e numerose ramificazioni arteriolari, fino al circolo
capillare, sono autosimili. Si viene a creare una vera e
propria ramificazione arboriforme che ad ogni
ingrandimento successivo ricorda la precedente scala di
grandezza.
I vasi retinici si suddividono e ramificano fino a
diventare così stretti che i globuli rossi, per passare,
sono costretti a disporsi in fila. Queste ramificazioni
diventano così numerose da raggiungere ed irrorare tutta
la retina. Per necessità fisiologica i vasi sanguigni
devono eseguire un pò di magia dimensionale per
comprimere una superficie immensa, di scambio tra sangue
e tessuti, in un volume limitato. Queste molteplici ed
autosimili diramazioni sono FRATTALI.
La struttura di questa circolazione ha consentito alla
natura di risolvere il problema in modo così efficiente
da permettere un perfetto e completo scambio nutrizionale
con la retina.
Con l'ausilio del calcolatore è possibile realizzare un
modello frattale dell'albero vascolare retinico, con
possibilità di ottenere successivi ingrandimenti
matematici fino alle più piccole ramificazioni.
Un modello di questo tipo è molto suggestivo e
realistico; è possibile modificare il decorso,
l'inclinazione, il tipo di ramificazione, l'angolo di
biforcazione ed il numero di successive divisioni. Con
queste modifiche possiamo rendere il modello più o meno
sovrapponibile ad una circolazione retinica normale o
renderlo "patologico" in modo da poter
quantizzare e misurare il grado di irregolarità.
E' possibile realizzare questi modelli frattali con le
funzioni iterative di Barnsley. Con l'immissione di pochi
valori numerici (parametri) si ottengono immagini
frattali molto realistiche della struttura ramificata
vasale. Modificando solo lievemente i suddetti parametri
è possibile modellare l'aspetto del frattale e renderlo
più o meno realistico.
Un risultato simile è possibile ottenerlo anche con
immagini frattali informatiche realizzate tracciando
graficamente segmenti ed applicando regole di
trasformazione un certo numero di volte.
Lo sviluppo embrionale dei vasi provenienti dalla
carotide interna verso la regione della coppa ottica
avviene per gemmazione di alcuni semplici tubi
endoteliali. Secondo alcuni Autori i vasi ed i capillari
retinici si sono formati , verso il quarto mese di
gestazione, da un processo di gemmazione dei vasi
presenti. Proprio per questa caratteristica gemmazione
nello sviluppo della circolazione retinica, l'aspetto
finale è suggestivamente simile ad un frattale
realizzato con aggregazione per diffusione. Questo si
sviluppa per gemmazione di ramificazioni che sono
determinate dall'aggregazione di punti che si muovono
casualmente.
Queste descrizioni di frattali possono essere utili solo
come strumento descrittivo e teorico, o come curiosità,
ma trovano scarso impiego pratico.
Nella descrizione ed interpretazione frattale della
circolazione retinica è necessario ipotizzare e
realizzare uno studio che ci permetta di quantizzare e
codificare alcuni parametri strutturali e morfologici
facilmente ripetibili, che forniscano una misura della
sua irregolarità.
La misura del grado di irregolarità di un modello
frattale è data dalla DIMENSIONE FRATTALE
("D"), che viene calcolata con la seguente
formula:
D = -Log N / Log 1/R
dove R è l'unita' di misura ridotta rispetto alla
precedente osservazione (esempio: unità di misura
ridotta a 1/3, R=3), N è il numero di unità R calcolate
nella misurazione della figura frattale presa in
considerazione (esempio: se R=3 cioè 1/3 di cm,
l'immagine frattale è lunga 3 cm , tenendo conto anche
delle irregolarità che possono essere misurate con la
suddette unità, N=9).
E' possibile calcolare la dimensione frattale D(dv) del
DECORSO VASALE RETINICO. Prendiamo in considerazione un
tratto vasale, più o meno irregolare secondo la
situazione patologica presente, della dimensione di
un'unità di misura arbitraria; tale unità di misura
viene ridotta ad un valore arbitrario (per esempio ad
1/5, R=5) della sua lunghezza, questo è il nostro regolo
di misura per calcolare la lunghezza del vaso preso in
considerazione, tenendo conto delle irregolarità che
possono essere misurate. In tal modo avremo il valore
della dimensione frattale D(dv) (con N=6 per esempio):
D(dv) = -Log 6 / Log 1/5= 1.113
Questo valore di D(dv) indica l'irregolarità del tratto
vasale preso in considerazione, nel nostro esempio la
dimensione frattale è lievemente superiore a 1 (un
oggetto lineare ha dimensione 1), quindi un grado di
irregolarità basso. Con i valori superiori di D(dv)
(fino a 2, il piano ha dimensione 2) aumenta
l'irregolarità. Un vaso molto tortuoso avrà
D(dv)=1,828, mentre un vaso retinico rettilineizzato per
trazione della limitante interna avrà D=1,012, molto
vicino a 1.
In questo modo è possibile quantizzare il grado di
irregolarità del decorso vasale.
Nella tabella 2.3 sono riportati i valori di D(dv)
calcolati in alcuni pazienti senza patologia vascolare
retinica, in coda alla stessa è stato calcolato il
valore medio.
La dimensione frattale può essere calcolata per un solo
tratto vasale, che a noi interessa, in tal caso possiamo
chiamarla segmentaria D(dv)s, oppure globale di tutta la
circolazione retinica D(dv)g. Può riguardare solo le
arterie (Dimensione arteriosa: D(dv)a) o solo le vene
(Dimensione venosa: D(dv)v):
D(dv) dimensione vasale frattale D(dv)s (dimensione
segmentaria) D(dv)g (dimensione globale ) D(dv)a
(dimensione arteriosa) D(dv)v (dimensione venosa)
Ovviamente esiste una evidente differenza tra Da e Dv
dovuta alla diversità di decorso dei vasi arteriosi e
venosi. Tale differenza può variare, aumentare o
diminuire, in base alle patologie vascolari del fondo.
Per calcolare la dimensione globale di decorso è
necessario eseguire una media delle varie dimensioni
arteriose e venose rilevate in più segmenti. Nella
tabella 2.4 sono riportati alcuni esempi dei valori di Da
e Dv in alcune situazioni patologiche.
Un altro valore importante è la differenza tra Dv e Da,
nel soggetto normale oscilla tra 0.005 e 0.050. Questo
valore indica che l'irregolarità di decorso è
lievemente maggiore nella circolazione venosa. Può
aumentare per riduzione di Da o per incremento di Dv.
Per esempio nella tortuosità vasale da trazione della
limitante interna (trazione sia dei vasi arteriosi che
venosi) il valore di Dv-Da è molto ridotto, quasi uguale
a 0, anche se i valori assoluti di Dv e Da sono aumentati
per maggiore irregolarità vasale complessiva.
Nella notevole tortuosità venosa di un emangioma la
differenza Dv- Da è molto aumentata perché Dv ha un
valore vicino a 2, quindi è un frattale vicino ad una
forma bidimensionale.
Studiando e confrontando con cura i vari valori parziali
e specifici di D è possibile catalogare e monitorizzare
con precisione il decorso dei vasi nelle varie patologie
della circolazione retinica.
In tal modo è possibile misurare il grado di
irregolarità con un parametro specifico e sensibile,
senza limitarsi solo a dare un giudizio soggettivo,
superficiale e non quantizzabile dei vasi retinici.
Ora esaminiamo complessivamente la distribuzione
ramificata dei vasi retinici. La circolazione retinica
complessiva può essere paragonata ad un modello frattale
realizzato per diffusione la cui dimensione frattale ha
valore di circa 1.71 (valori variabili tra 1.66 e 1.76)
.Questo valore indica l'irregolarità, comprendendo
anche le biforcazioni e la diffusione su tutto il piano
retinico (quasi come il piano a due dimensioni) della
distribuzione dei vasi.
Il valore di D è maggiore rispetto al valore di D(dv)
perché quest'ultimo indica solo l'irregolarità del
decorso lineare (lievemente superiore ad una dimensione)
del vaso e non l'irregolarità di distribuzione
determinata dalle diramazioni nel piano.

13 - IL FLUSSO FLUORESCEINICO VENOSO
Dopo l'iniezione della fluoresceina per via venosa,
durante l'angiografia retinica a fluorescenza, è
possibile evidenziare nella circolazione retinica un
particolare tipo di flusso del colorante.
La fluoresceina penetra nelle vene lungo le pareti
formando un cilindro di colorante. Questa colonna di
colorante è a forma di lamina addossata alla parete.
Ogni corrente venosa che si getta in un tronco più
grande rimane ben distinta, conservando la sua
indipendenza (Fig.
2.23, 2.24). Questo aspetto viene conservato per più
confluenze fino a mostrarsi con caratteristiche
multilaminari nei tronchi più grandi, e successivamente
non è più individuabile.
Questo aspetto multilaminare, risultante dalla confluenza
di più colonne fluoresceiniche di capillari e di venule,
non è sempre visibile per l'uniformità di contrasto
delle lamine.
Esaminando queste numerose lamine a ingrandimenti sempre
maggiori, eventualmente con accorgimenti che ne
permettano l'individuazione, vengono rivelati dettagli
sempre maggiori. Ogni lamina è formata da numerose altre
lamine più piccole, provenienti dalla confluenza di vasi
minori, queste ultime, ad ingrandimenti ulteriori, sono
composte da altre lamine minori originate da vasi più
piccoli.
Questa organizzazione laminare presenta una
autosomiglianza a più scale, con caratteristiche
aleatorie, e quindi di tipo frattale.
L'aspetto laminare della fluoresceina è simile ai
dettagli infinitesimali rivelati dall'ingrandimento
dell'attrattore di Hènon e agli allungamenti e
ripiegamenti caratteristici del mescolamento dei fluidi.
Questo moto regolare laminare è caratteristico di un
fluido che scorre tranquillamente senza l'intervento di
forze esterne o anomalie delle pareti. Al contrario,
quando un fluido è messo in agitazione da una
irregolarità della parete o da anomalie vasali, questo
tranquillo e ordinato comportamento diventa turbolento.
Non è possibile più separare le lamine, queste si
aggrovigliano nei modi più contorti. In questi sistemi
si formano gorghi, vortici e spirali di ogni tipo. Si
viene a creare una vera turbolenza, con un regime
caotico.
Tutti i sistemi con comportamento turbolento son
"dispersivi", cioè disperdono energia, per
l'attrito del flusso idrodinamico. Una evidente
conseguenza dell'attrito è che il moto tende ad
arrestarsi a meno che non venga immessa energia.
Il rilevamento, con immagini digitalizzate ed elaborate
al computer, del movimento, della qualità delle lamine e
della eventuale comparsa di turbolenze può fornire
informazioni molto utili sulla qualità della
circolazione retinica e sull'origine delle anomalie.
L'elaborazione e lo studio di questi dati potrebbe
consentire la progettazione di sofisticati sistemi di
riconoscimento delle forme, che siano in grado di
rilevare le minime variazioni della regolarità del
flusso con la possibilità di una identificazione delle
anomalie vasali.
14 - LE LESIONI RETINICHE
La distribuzione e la concentrazione delle lesioni
retiniche (come emorragie, essudati, microaneurismi,
ecc.), presenti in numerose patologie retiniche, sono
spesso oggetto di studio, con tentativi di
quantizzazione. Ciò molto spesso non è possibile per la
natura aleatoria dei fattori che intervengono.
Servendosi del concetto di Polvere di Cantor è possibile
calcolare la distribuzione delle suddette lesioni nella
retina. Anche se il modello è una funzione e non impiega
dati reali, esso ricorda la distribuzione delle
emorragie, degli essudati, dei microaneurismi e di molte
altre lesioni, che altrimenti risulterebbe indefinibile.
Una immagine di distribuzione simile, casuale e con
caratteristiche frattali come la polvere di Cantor, è
determinata dal movimento browniano per generare un
"universo" disseminando un punto ad ogni sosta;
" D " (dimensione frattale) rappresenta il
grado di ammassamento medio dei punti.
I frattali basati sull'insieme di Cantor non descrivono
soltanto il modo in cui tali lesioni si raggruppano e si
distribuiscono sulla retina, ma anche come si manifestano
nel tempo.
Questo metodo descrittivo comporta una gerarchizzazione
degli ammassi a più livelli. E' possibile incontrare
ammassi di lesioni che a loro volta sono formati da altri
di natura simile ma di dimensioni minori. In base alla
concentrazione ed alla distribuzione sarà possibile
calcolare una dimensione frattale " D ". In tal
modo si quantizzerà con un unico valore, tipico della
lesione, la distribuzione spaziale.
Nelle figure 2 e 3 sono riportati alcuni esempi di
lesioni del fondo che presentano le caratteristiche di
distribuzione gerarchica, con omotetia a più scale,
legata al caso. Per esempio la distribuzione di Drusen
retiniche è organizzata in raggruppamenti grandi che a
loro volta sono formati da altri più piccoli. Una
organizzazione simile è presente anche nelle lesioni
emorragiche delle occlusioni venose, anch'esse formate da
raggruppamenti di altre più piccole .
Questo modello frattale è un tentativo di evitare
l'impreciso ed indaginoso metodo di conteggio e di
descrizione incompleta delle lesioni retiniche, in modo
di poter monitorizzare con maggior precisione ed
obiettività l'evoluzione della patologia.
15 - LE CELLULE NERVOSE RETINICHE
Le cellule nervose sottostanti alle cellule visive si
trovano negli strati più interni della retina e sono
connesse con le cellule recettrici visive. Esistono più
tipi di cellule nervose retiniche (bipolari, multipolari,
amacrine, orizzontali) con varie funzioni di conduzione e
di interconnessione; dello stesso tipo si possono trovare
numerose variazioni morfologiche.
Queste cellule nervose, come tutti i neuroni, presentano
una caratteristica forma, anche se variabile tra i vari
tipi, con prolungamenti di aspetto ramificato, detto
dendrite. Questi dendriti si ramificano più volte dando
luogo ad una caratteristica forma frattale autosimile a
più scale.
Se si esaminano tali cellule a basso ingrandimento, si
possono osservare ramificazioni asimmetriche connesse con
i corpi cellulari.
A ingrandimento leggermente superiore si osservano
ramificazioni più piccole a partire da quelle più
grandi. Aumentando ulteriormente l'ingrandimento, si può
scorgere un altro livello di dettaglio: ramificazioni di
ramificazioni di ramificazioni.
Di fronte a due fotografie di dendriti a differenti
livelli di ingrandimento (senza alcun riferimento di
scale sull'immagine), può essere difficile riconoscere a
quale ingrandimento corrisponda una determinata
fotografia.
Nelle figure sono riportati alcuni modelli di cellule
nervose retiniche realizzati con il computer.
Le cellule amacrine (particolare dell'Insieme di
Mandelbroit) sono piriformi e grosse (A), i loro
prolungamenti emanano dalla estremità inferiore della
cellula ed hanno i caratteri di dendriti (B), non è
dimostrabile il neurite.
Le cellule bipolari ( particolare dell'Insieme di
Mandelbroit) hanno il corpo ovoidale (A), da ciascun polo
parte un prolungamento, quello ascendente è considerato
come un dendrite (B) e si risolve con un pennacchio (C),
il prolungamento discendente (D), considerato un neurite,
con la sua arborizzazione si mette in rapporto con le
cellule multipolari.
Le cellule orizzontali (A) ( particolare dell'Insieme di
Julia) hanno un neurite grosso e lungo con
un'arborizzazione complicata ed estesa (B), il dendrite
può essere corto (C).
16 - ORGANIZZAZIONE RETINICA PREGANGLIARE FUNZIONI DI
CONTROLLO DELL'IMMAGINE.
Le vie retiniche di conduzione del messaggio visivo
costituiscono una rete molto complessa. Le strutture
cellulari (cellule orizzontali, cellule amacrine, e
interneuroni) che compongono questi canali retinici
presentano alcune funzioni altamente specializzate che
permettono una selezione, una rielaborazione ed un
miglioramento qualitativo dell'immagine trasmessa.
La principale di queste funzioni è quella di CONTROLLO
RETROATTIVO. Con questo meccanismo le rete elaborativa
pregangliare riesce a correggere gli "errori",
che si producono a livello recettoriale, attraverso il
caos deterministico.
Un sistema retroattivo è caratterizzato da una
circolazione dell'informazione prodotta, che viene
ripresentata all'ingresso e rielaborata per poi essere
nuovamente ripresentata all'ingresso. Questa retroazione
(iterazione) può avvenire un numero infinito di volte.
Un circuito retroattivo con un elemento non lineare
presenterà un risultato finale modificato; l'immagine
finale dipende dall'elemento non lineare. Questi sistemi
non lineari presentano comportamenti di notevole
complessità, cioè di tipo caotico.
Consideriamo ora un sistema contenente un meccanismo non
lineare, i punti dell'immagine potranno essere accoppiati
in due modi: Accoppiamento diffusivo o Accoppiamento
sottrattivo. La diffusione dovuta ad una sfuocatura di
due punti dell'immagine porta ad una sovrapposizione e
perciò all'accoppiamento di punti vicini dell'immagine;
le intensità si sommano, cioè ciascun punto riceve un
"rinforzo" dai punti vicini, si ha così un
accoppiamento diffusivo.
Secondo l'accoppiamento sottrattivo, le intensità dei
punti vicini non vengono sommate ma sottratte, come
avviene nelle cellule nervose della retina.
L'accoppiamento dell'immagine è importante per spiegare
come nasca l'ordine in un sistema a retroazione.
Prendiamo un sistema retroattivo e facciamolo partire da
una immagine arbitraria, dopo un centinaio di iterazioni
l'immagine non si modifica più, il sistema dinamico non
lineare ha raggiunto una immagine stabile, un punto
fisso. Con questa immagine ottenuta si è raggiunto
l'ordine con forme stabili. Con un accoppiamento diverso,
ma con la stessa immagine emerge un punto fisso diverso.
Anche l'immagine di partenza è determinante per
l'immagine finale.
Dopo molte iterazioni, il punto fisso di un sistema
dinamico non lineare rappresenta il punto di quiete o
immagine stabile. Lo stesso punto fisso, immagine finale,
a cui si arriva dalle diverse immagini di partenza, viene
chiamato attrattore. L'insieme delle immagini di partenza
che porta allo stesso punto fisso forma il bacino di
attrazione dell'attrattore. Di conseguenza due immagini
lievemente diverse, appartenenti allo stesso bacino di
attrazione di uno stesso attrattore, dovrebbero evolvere
verso lo stesso punto fisso, cioè la stessa immagine
finale.
Una immagine lievemente alterata dovrebbe evolvere verso
la stessa immagine finale di una originariamente
perfetta. Se facciamo circolare un'immagine distorta nel
sistema di retroazione non lineare, la distorsione
scompare e si raggiunge lo stesso punto fisso
dell'immagine originale non alterata.
Questi sistemi capaci di ripristinare perfettamente
l'integrità di un'immagine, partendo da informazioni
incomplete, si chiamano memorie associative. Quindi il
circuito a retroazione presenta questa proprietà.
Anche nella retina si trovano elementi che costituiscono
un sistema simile di collegamenti e che, attraverso
elementi di accoppiamento non lineari sottrattivi,
retroagiscono sui conduttori d'ingresso di altri
dispositivi non lineari; le caratteristiche dei singoli
elementi di accoppiamento determinano il punto fisso
raggiunto.
Le cellule nervose retiniche che contengono le non
linearità hanno conduttori d'ingresso, che sono i
dendriti, mentre le uscite, che sono gli assoni,
trasmettono i segnali alle sinapsi, che rappresentano gli
elementi di accoppiamento con altre cellule nervose. In
questo sistema sono possibili innumerevoli accoppiamenti.
In pratica il sistema a retroazione ha la possibilità di
ricostruire in modo associativo un'immagine a partire da
una incompleta. Con questo meccanismo possono essere
ripristinate immagini di partenza sfuocate, disturbate o
incomplete.
Se l'immagine di partenza è estremamente alterata, non
appartiene più al bacino di attrazione del punto fisso
dell'immagine da ripristinare, quindi le iterazioni
portano ad un altro punto fisso: una immagine diversa.
E' possibile intuire che a livello delle interconnessioni
pregangliari retiniche è presente un sistema molto
complesso di retroazione non lineare, che permette già a
questo livello una elaborazione e una depurazione
dell'immagine, o di parte di essa.
L'immagine presente a livello recettoriale viene, con
questo sistema, elaborata e migliorata nella rete delle
interconnessioni pregangliari. Quindi ai fini del
risultato funzionale può risultare più grave un danno
alle interconnessioni , che riduce o sopprime questo
meccanismo di memoria associativa, piuttosto che una
lesione parziale dei recettori. L'immagine presente a
livello recettoriale è solo la partenza del circuito
retroattivo, le eventuali distorsioni o difetti possono
essere ripristinati dal suddetto meccanismo.
17 - LA FOTOCHIMICA DELLA VISIONE
La rodopsina è il pigmento contenuto in maniera
altamente organizzato nell'articolo esterno dei
fotorecettori. Il processo fondamentale dell'assorbimento
della luce da parte della rodopsina consiste nella
trasformazione del retinale 11-cis in tutto-trans,
prodotto molto instabile, attraverso varie tappe. Una
serie di reazioni chimiche porta ad una reisomerizzazione
, che rigenera la rodopsina in forma 11-cis. La
trasformazione da 11-cis a tutto-trans determina lo
sbiancamento della rodopsina con una iperpolarizzazione
della cellula ed una modificazione della resistenza
elettrica della membrana.
Questa continua reazione chimica bidirezionale è
ovviamente influenzata, o meglio determinata, dalla
quantità e dalla qualità della luce che arriva alla
retina. Per l'estrema variabilità dello stimolo
luminoso, in ambiente di vita normale, la reazione
chimica suddetta presenta caratteristiche aperiodiche, o
meglio caotiche, influenzata anche da fattori endogeni
elettrochimici.
In questa reazione le concentrazioni dei componenti non
raggiungono valori costanti e neppure oscillano
periodicamente, ma piuttosto aumentano e diminuiscono in
modo apparentemente casuale e imprevedibile. Si viene a
creare il "CAOS CHIMICO", come in molte altre
reazioni chimiche strettamente dipendenti da fattori
esterni variabili in modo aleatorio.
E' possibile rappresentare anche questa reazione, come
tutti i sistemi dinamici con carattere caotico, con un
"attrattore strano" nello spazio delle fasi.
Ad una reazione chimica di questo tipo corrisponderà un
ben caratteristico potenziale elettrico, che per un
singolo stimolo luminoso sarà deterministico e ben
codificabile, perché dipendente da una unidirezionalità
del sistema dinamico. Le caratteristiche elettriche
"in ambiente" saranno del tutto aleatorie per
comparsa delle caratteristiche caotiche della reazione.
Sarebbe certamente molto interessante studiare le
caratteristiche di un tracciato elettrico di questo tipo
nello spazio delle fasi.
In tal modo i risultati non sarebbero influenzati dalle
condizioni di esecuzione e di adattamento, ma
dipenderebbero solo in condizioni fisiologiche, dalle
caratteristiche caotiche esogene ed endogene.
Secondo alcuni autori il caos chimico non rappresenta
niente di più che fluttuazioni incontrollate che
spingono a caso il sistema.
Altri autori propongono che il caos chimico sia un
fenomeno genuino inerente alla dinamica di alcuni sistemi
chimici oscillanti.
18 - LA COROIDE
La coroide è formata sostanzialmente da vasi, arteriosi
e venosi, distribuiti in tre strati in base alle loro
dimensioni. I primi due strati vascolari sono nella
lamina vascolosa, in quello più superficiale decorrono i
grossi vasi, mentre in quello più profondo i vasi più
piccoli. Il terzo strato vascolare è la coriocapillare,
formata da vasi capillari riuniti a rete.
Questa distribuzione in più strati, in ciascuno si
trovano vasi simili ma più piccoli dello strato
precedente, presenta caratteristiche di autosomiglianza
frattale. I vasi arteriosi della coroide presentano
ramificazioni di tipo frattale; i vasi principali, nello
strato più superficiale, si ramificano in una serie di
vasi più piccoli, che decorrono nello strato più
profondo, questi si ramificano, a loro volta, in vasi di
calibro ancora minore e poi in capillari.
La coriocapillare, a sua volta, ha una sua struttura
autosimile per l'organizzazione a lobuli; non è un
sistema liberamente anastomizzato, come si riteneva in
passato, ma è diviso in lobuli indipendenti, in maniera
simile a quella del fegato.
Queste strutture elementari, i lobuli, sono delimitate
dai capillari più grossi orientati circonferenzialmente;
all'interno dei lobuli si trovano i capillari più
piccoli che delimitano aree di tessuto contenenti
capillari ancora più piccoli.
I capillari scaricano il sangue nelle vene minori, che si
riuniscono a "vortice" nelle vene più grandi,
fino alle vene vorticose.
In tal modo la circolazione venosa viene ad assumere
l'aspetto di un insieme di vortici di varie dimensioni.
Questo struttura vorticosa ha radici matematiche, è
simile alle numerose immagini frattali, anch'esse a
vortice, di alcuni particolari dell'Insieme di
Mandelbroit, o ad alcune strutture realizzabili al
computer con orbite sovrapposte generate da equazioni
matematiche complesse contenenti funzioni
trigonometriche.
19 - IL NERVO OTTICO
Il nervo ottico è formato di fibre nervose midollate,
senza nevrilemma, che si raggruppano in fascetti detti
"fascetti primari".
Questi a loro volta si riuniscono a formare i fasci, che
sarebbero circa un migliaio.
Questi livelli di raggruppamento di fibre nervose sono
determinati dalla nevroglia. Uno straterello di nevroglia
è alla superficie dell'asse connettivale che accoglie i
vasi centrali della retina. A ciascun fascio di fibre la
nevroglia forma un manicotto, da questo emanano sepimenti
nevroglici che penetrano nel fascio e lo dividono in
fascetti più piccoli (fascetti primari).
Questa organizzazione strutturale in fasci sempre più
piccoli è di tipo frattale. Anche se non esiste
un'autosomiglianza completa tra i vari livelli di fasci
di fibre, è possibile certamente notare una somiglianza
organizzativa e dei setti di nevroglia.
L'autosomiglianza a un certo livello si ferma, però i
dettagli dei fascetti primari sono simili, seppur non
necessariamnente identici, a quelli della struttura vista
a ingrandimento minore, cioè ai fasci.
20 - I MOVIMENTI OCULARI DI FISSAZIONE
Nella fissazione di un oggetto su uno sfondo uniforme,
l'occhio presenta una serie di micromovimenti, necessari
per rendere possibile la continuità della visione.
Questi movimenti, i soli che abbiano luogo durante la
pausa di fissazione, sono sufficienti a mantenere la
visione anche per tempi prolungati.
Ad un primo esame superficiale della registrazione di
questi movimenti durante la fissazione, sembrano
oscillazioni apparentemente casuali. Esaminando
attentamente le componenti a varie scale temporali,
possiamo notare alcune caratteristiche importanti. Ad una
prima scala di minuti riscontriamo i "MOVIMENTI DI
DERIVA" (Drift), movimenti lenti e a bassa
frequenza; ad un successivo ingrandimento, scala di
secondi, notiamo la presenza di piccoli movimenti a
scosse, i "MICROSACCADI.
Esaminando ulteriormente il tracciato, ad una scala
minore, è possibile osservare un tremore ad alta
frequenza (70-90 oscillazioni al secondo).
Questi movimenti sono autosimili a scale temporali
diverse; le leggi che regolano questa autosomiglianza non
sono lineari, ma aleatorie.
Queste osservazioni suggeriscono che i sistemi di
controllo di questi movimenti sono di tipo caotico, in
una situazione fisiologica.
In una disfunzione a carico dei sistemi preposti alla
stabilizzazione della fissazione si ha la comparsa di
nistagmo patologico, movimenti ritmici con
caratteristiche di sistema dinamico ciclico.
Analizziamo la rappresentazione dello spazio delle fasi
per i movimenti oculari di fissazione, in cui gli assi
sono rispettivamente la posizione e la velocità. Il
comportamento della dinamica dei movimenti di fissazione
fisiologici è del tipo ATTRATTORE STRANO; mentre in caso
di nistagmo, che può essere paragonato ad un sistema
periodico, l'attrattore è a CICLO LIMITE. In condizioni
estreme di notevole riduzione dei movimenti, per
incapacità di fissazione o per anomalie della motilità
estrinseca, l'attrattore tende a diventare a PUNTO FISSO.
L'attrattore strano, fisiologico, è espressione di un
sistema dinamico caotico, su cui agiscono numerose
componenti di tipo neurogeno, psicofisico e meccanico. Un
sistema di movimenti di fissazione necessita proprio di
condizioni caotiche per risultare efficace e rapido,
rispondendo alle necessità di mantenimento della
visione, senza permettere la degradazione dell'immagine
per una eccessiva persistenza nella stessa area retinica.
L'andamento periodico del nistagmo patologico non
permette una distribuzione caotica dello stimolo
luminoso, ma, con i suoi movimenti ritmici, stabilizza
l'immagine sempre sulle stesse aree retiniche; in tal
modo l'immagine degrada e determina una riduzione della
capacità visiva. Quindi la riduzione funzionale nel
nistagmo non sarebbe dovuta alla incapacità di catturare
l'immagine, per il movimento rapido dell'occhio,
piuttosto al fatto che non vengono stimolate aree
retiniche con distribuzione di tipo aleatoria, ma di tipo
deterministico.
In pratica anche nella dinamica della fissazione la
fisiologia si associa alle leggi del caos, mentre la
patologia mostra una periodicità che impedisce al
sistema di rispondere alle esigenze funzionali della
biochimica retinica e a quelle ottiche.
21 - I FOSFENI
Le immagini luminose che talvolta vediamo apparire dietro
le palpebre chiuse o nell'oscurità completa sono dette
fosfeni (dal greco phos=luce e phanein=apparire) e
derivano dall'autostimolazione delle fibre ottiche.
Possono essere provocate comprimendo i bulbi oculari,
dall'assunzione di alcune droghe o da alcune situazioni
patologiche sistemiche o del sistema nervoso centrale.
La forma ed i colori dei fosfeni sono molteplici,
dipendono dalle situazioni che li determinano e da
componenti individuali. Le immagini più caratteristiche
e frequenti sono: a "fortificazione", a
"spirale", a cerchi concentrici, a dischi
colorati, a "onde", e con tante altre svariate
forme.
Praticamente tutte queste forme di fosfeni sono di
aspetto frattale, con caratteristiche autosimili a più
scale, e facilmente riproducibili con l'elaboratore,
utilizzando algoritmi matematici produttori di frattali.
Questi fosfeni sono frattali perché sono generati nelle
vie ottiche da meccanismi a catena automoltiplicanti,
simili ai procedimenti iterativi generatori di frattali.
I fosfeni spontanei rappresentano per le vie ottiche,
come avviene in tutte le linee di trasmissione di dati,
quelle inevitabili e numerose fluttuazioni spontanee che
possiamo chiamare "rumori".
La distribuzione nel tempo del "rumore" ha
caratteristiche molto curiose e molto importanti dal
punto di vista concettuale e pratico.
Analizzando nel tempo la comparsa di questi
"errori", nella conduzione dello stimolo
luminoso, in modo via via più fine, è possibile notare
che in alcune ore non c'è alcun "errore". In
altri momenti compaiono "raffiche di errori",
che nel loro ambito presentano diversi momenti di quiete
di durata minore. Analogalmente tra questi momenti di
quiete compaiono nuove "raffiche di errori"
più brevi delle precedenti. La distribuzione di ciascun
ordine di raffiche è identica, dal punto di vista
statistico, rispetto all'ordine immediatamente superiore.
Si scopre così un nuovo esempio di omotetia interna, con
una dimensione frattale.
Un modello valido per le raffiche di errori è una
variante aleatoria dell'insieme di Cantor (vedi parte I),
denominata "Polvere di Levy".
Questo comportamento aleatorio della comparsa dei fosfeni
dipende dalle numerose componenti esogene ed endogene con
caratteri caotici.
Come in tutti i sistemi caotici, una causa piccolissima
che sfugga alla nostra attenzione determina un effetto
considerevole con possibile scatenamento di una crisi con
"raffiche di fosfeni". Quindi una piccola
differenza può portare a una notevole variazione di
comportamento, che dal punto di vista matematico si
chiama "biforcazione".
A questo punto ci si chiede se il passaggio dal
comportamento normale a quello di crisi di fosfeni non
sia dovuto a una biforcazione.
Se così fosse, le terapi

home page